

细菌细胞分裂相关研究:
[重要进展]FtsZ原丝纤维是如何进一步组装成Z-环?回答这个未解之谜的关键之一是要知道FtsZ原丝纤维之间是否存在相互作用。我们在晶体结构中观察到FtsZ原丝纤维之间的相互作用面,进一步通过和北京大学昌增益实验室合作,我们以定点插入非天然氨基酸后进行光交联实验的方法证明了我们在体外观察到的相互作用面在细胞中真是存在,如果这个相互作用面被彻底破坏则导致细胞不能分裂。(关凤慧等,eLife,2018)[文章链接]。
[重要进展] 细菌细胞分裂研究中的关键问题之一是:Z环是如何将GTP水解的化学能转化为向内收缩的张力?为了回答这个问题,我们解析了FtsZ原丝纤维在弯曲状态下的结构,并根据这个结构设计了一系列突变,通过在大肠杆菌上进行的体内互补实验,同时结合GTPase酶活测定、电子显微镜负染观测,辅以分子动力学模拟,提出并验证了Z-环是如何将GTP水解的化学能转换为原丝纤维的构象变化的分子机制(李颖等,Science,2013)[文章链接]
离子通道相关研究:
[重要进展]我们与永利yl23411官网登录冷冻电镜中心联合解析了人源a5b3 GABAA受体结构,这是永利yl23411官网登录第一个以单颗粒冷冻电镜技术得到的蛋白质原子分辨率结构。GABAA受体是中枢神经系统中介导抑制性突触传递的氯离子通道,是许多抗抑郁症药物和麻醉药物的靶蛋白。我们揭示了异源五聚a5b3 GABAA受体特殊的亚基组装形式,以及γ-氨基丁酸(GABA)激活GABAA氯离子通道开放的机制。(刘斯等,Cell Res,2018)
[重要进展]通过结合X-射线晶体学和电生理学来研究钾离子通道的钡离子阻断机制。我们解释了钾离子对于钡离子从钾离子通道逃逸存在的三个效应:外部锁定效应、增强效应和内部锁定效应,提出这三个效应是由于能量平衡被破坏而导致的(郭蕊等,J Gen Physiol,2014)[文章链接]。
[重要进展]同UTSW姜有星课题组合作,我们以MthK通道为模型,首次对离子通道领域近二十年未解的一个现象:反常摩尔混合效应提出了一个合理的机制,即离子在穿过离子通道过程中的能量平衡被破坏,同时提出了钾离子通道选择性通透钾离子的机制(叶升等,Nat Struc Mol Biol,2010)。
1. Z环收缩的分子机制
原核细胞的骨架蛋白FtsZ(细胞分裂温度敏感突变体Z)能够自组装成为细菌细胞分裂过程中的一个关键细胞器―Z环,细胞分裂时,随着分裂隔膜形成,Z环将水解GTP释放的化学能转化成为向内拉伸的机械力,缩小自身直径,拉动隔膜内陷,直至最终收缩分裂细胞。围绕细菌细胞骨架蛋白FtsZ研究领域中的一个关键问题:Z环是怎样将GTP水解的化学能转换为向内收缩的张力?我们根据所得到的FtsZ原丝纤维的晶体结构,设计一系列的FtsZ突变体,直接在大肠杆菌上进行体内互补实验、同时结合体外GTPase酶活测定、电子显微镜负染观测,辅以分子动力学模拟,提出并验证了Z环是怎样将GTP水解的化学能转换为向内收缩的张力的分子机制(见图1、2)(Li et al, 2013)
 |
| 图1: FtsZ单体在结合GTP和结合GDP时的主要构象变化发生在T3环状结构区(T3 loop),GTP的g-磷酸基团能够和T3环的主链胺基形成两个强氢键相互作用,从而锁定T3环处于一种紧密结合构象(T 构象)。当FtsZ结合GDP,失去了这两个强氢键相互作用后,T3环除了可以处于紧密结合构象之外,还可以处于一种松弛构象(R 构象)。此外,在FtsZ原丝纤维的垂直构象中有两个非常关键的结构特征,首先,T3环处于紧密结合构象并和上面亚基的T7环相互作用,使得T7环上两个保守的天冬氨酸靠近GTP g-磷酸基团附近的一个水分子,在催化GTP水解的反应中,这两个天冬氨酸的酸性残基极化这个水分子,使其能够进攻GTP的 g-磷酸基团。其次,在FtsZ原丝纤维的垂直构象中,相邻两个FtsZ亚基紧密结合,严丝合缝,夹在两亚基之间的GDP是处于一种封闭状态,除非两个亚基打开,是无法自由出来。因此我们推测,当GTP发生水解,GTP g-磷酸基团和b-磷酸基团之间的共价键断裂,由于二者都携带负电荷,它们之间会产生一个巨大的排斥力,当二者被封闭在两个FtsZ亚基之间时,就会如图所示,这个巨大的排斥力驱使GTP g-磷酸基团推动T3环发生一个从紧密结合构象到松弛构象的构象变化,从而使两个FtsZ亚基之间的结合从T3环和T7环接触处打开,相邻亚基围绕支点发生折叠。GTP水解的化学能就这样转变成为相邻FtsZ亚基之间的构象变化。 |
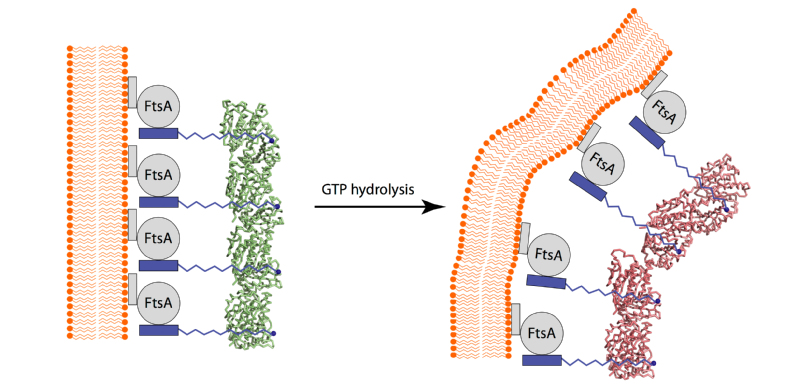 |
| 图2:上述GTP水解而导致的FtsZ原丝纤维构象变化如何转化为细菌细胞分裂向内收缩的张力呢?FtsZ的C端肽链没有一个稳定的构象,这段肽段会和FtsA相结合,而FtsA通过它的C端一个疏水的a-螺旋嵌在细胞膜上,Z环就是这样被固定在细胞膜上位于隔膜最内端(左)。上述GTP水解而导致FtsZ原丝纤维的构象变化则通过FtsA施加一个改变细胞膜曲度的一个向内收缩的张力(右)。 |
为什么钾离子通道选择性地通透钾离子,而不通透直径比钾离子更小的钠离子?许多离子通道专家在试图回答这个问题时观察到钾离子通道存在一个生物物理现象-反常摩尔混合效应(Anomalous Mole-Fraction Effect)。2003年诺贝尔化学奖获得者MacKinnon通过对钾离子通道KcsA的研究发现,当钠离子浓度升高而钾离子浓度降低到3mM以下时,KcsA通道的离子筛结构塌陷为不通透构陷,因此MacKinnon认为离子筛结构的构象变化使得钾离子通道选择性地通透钾,然而这无法解释反常摩尔混合效应,我们在研究Mthk通道是发现:Mthk通道在极端条件下(钾离子完全不存在)是可以通透钠离子的,在此基础上加入微量(1mM)的钾离子则阻断钠电流,MthK这个性质和KcsA通道完全不同,反而和前述的其他离子通道专家所观察到实验现象类似。微量深入理解MthK的离子选择性质,我们结晶得到1.45埃分辨率的MthK通道。在这个高分辨率结构的基础上,我们看到在完全没有钾离子存在的条件下,MthK通道的离子筛维持通透构象;在高钾离子浓度条件下,钾离子在离子筛四个结合位点均衡分布;在低钾离子浓度条件下,钾离子在离子筛内倾向结合1,3位点,我们的结构分析和功能试验完全相符,解释了困扰离子通道专家近二十年的生物物理现象(Ye et al, 2010)